Citation: Junli Liu. Threshold dynamics of a time-delayed hantavirus infection model in periodic environments[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4758-4776. doi: 10.3934/mbe.2019239

| [1] | J. A. Reinoso and F. J. de la Rubia, Stage-dependent model for Hantavirus infection: The effect of the initial infection-free period, Phys. Rev. E, 87 (2013), 042706. |
| [2] | C. H. Calisher, W. Sweeney, J. N. Mills, et al., Natural history of Sin Nombre virus in western Colorado, Emerg. Infect. Dis., 5 (1999), 126–134. |
| [3] | J. N. Mills, T. G. Ksiazek, C. J. Peters, et al., Long-term studies of hantavirus reservoir populations in the southwestern United States: a synthesis, Emerg. Infect. Dis., 5 (1999), 135–142. |
| [4] | J. N. Mills, T. G. Ksiazek, B. A. Ellis, et al., Patterns of association with host and habitat: antibody reactive with Sin Nombre virus in small mammals in the major biotic communities of the southwestern United States, Am. J. Trop. Med. Hyg., 56 (1997), 273–284. |
| [5] | G. E. Glass, W. Livingston, J. N. Mills, et al., Black Creek Canal Virus infection in Sigmodon hispidus in southern Florida, Am. J. Trop. Med. Hyg., 59 (1998), 699–703. |
| [6] | G. Abramson and V. M. Kenkre, Spatiotemporal patterns in the Hantavirus infection, Phys. Rev. E, 66 (2002), 011912. |
| [7] | G. Abramson, V. M. Kenkre, T. L. Yates, et al., Traveling Waves of Infection in the Hantavirus Epidemics, Bull. Math. Biol., 65 (2003), 519–534. |
| [8] | L. J. S. Allen, M. Langlais and C. J. Phillips, The dynamics of two viral infections in a single host population with applications to hantavirus, Math. Biosci., 186 (2003), 191–217. |
| [9] | V. M. Kenkre, L. Giuggioli, G. Abramson, et al., Theory of hantavirus infection spread incorpo-rating localized adult and itinerant juvenile mice, Eur. Phys. J. B, 55 (2007), 461–470. |
| [10] | T. Gedeon, C. Bodelón and A. Kuenzi, Hantavirus Transmission in Sylvan and Peridomestic Environments, Bull. Math. Biol., 72 (2010), 541–564. |
| [11] | F. Sauvage, M. Langlais, N. G. Yoccoz, et al., Modelling hantavirus in fluctuating populations of bank voles: the role of indirect transmission on virus persistence, Journal of Animal Ecology, 72 (2003), 1–13. |
| [12] | C. Wolf, A mathematical model for the propagation of a hantavirus in structured populations, Discrete Contin. Dyn. Syst. Ser. B, 4 (2004), 1065–1089. |
| [13] | C. Wolf, M. Langlais, F. Sauvage, et al., A multi-patch epidemic model with periodic demography, direct and indirect transmission and variable maturation rate, Math. Popul. Stud., 13 (2006), 153–177. |
| [14] | J. A. Reinoso and F. J. de la Rubia, Spatial spread of the Hantavirus infection, Phys. Rev. E, 91 (2015), 032703. |
| [15] | J. Buceta, C. Escudero, F. J. de la Rubia, et al., Outbreaks of Hantavirus induced by seasonality, Phys. Rev. E, 69 (2004), 021906. |
| [16] | L. J. S. Allen, R. K. McCormack and C. B. Jonsson, Mathematical models for hantavirus Infection in rodents, Bull. Math. Biol., 68 (2006), 511–524. |
| [17] | R. Bürger, G. Chowell, E. Gavilán, et al., Numerical solution of a spatio-temporal gender-structured model for hantavirus infection in rodents, Math. Biosci. Eng., 15 (2018), 95–123. |
| [18] | R. Ostfeld and F. Keesing, Pulsed resources and community dynamics of consumers in terrestrial ecosystems, Trends in Ecology and Evolution, 15 (2000), 232–237. |
| [19] | K. D. Abbott, T. G. Ksiazek and J. N. Mills, Long-term hantavirus persistency in rodent popula-tions in central Arizona, Emerg. Infect. Dis., 5 (1999), 102–112. |
| [20] | A. J. Kuenzi, M. L. Morrison, D. E. Swann, et al., A longitudinal study of Sin Nombre virus prevalence in rodents in southwestern Arizona, Emerg. Infect. Dis., 5 (1999), 113–117. |
| [21] | J. K. Hale and S. M. Verduyn Lunel, Introduction to Functional Differential Equations, Springer, New York, 1993. |
| [22] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems,Math. Surveys Monogr. 41, AMS, Providence, RI, 1995. |
| [23] | X.-Q. Zhao, Basic reproduction ratios for periodic compartmental models with time delay, J. Dyn. Diff. Equat., 29 (2017), 67–82. |
| [24] | W. Walter,On strongly monotone flows, Ann. Polon. Math., 66 (1997), 269–274. |
| [25] | P. Magal and X.-Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM. J. Math. Anal., 37 (2005), 251–275. |
| [26] | X. Liang and X.-Q. Zhao, Asymptotic speeds of spread and traveling waves for monotone semi-flows with applications, Comm. Pure Appl. Math., 60 (2007), 1–40. |
| [27] | X. Wang and X.-Q. Zhao, Dynamics of a time-delayed Lyme disease model with seasonality, SIAM J. Appl. Dyn. Syst., 16 (2017), 853–881. |
| [28] | X.-Q. Zhao, Dynamical Systems in Population Biology, Springer-Verlag, New York, 2003. |
| [29] | Y. Yuan and X.-Q. Zhao, Global stability for non-monotone delay equations (with application to a model of blood cell production), J. Differ. Equations, 252 (2012), 2189–2209. |
| [30] | D. Posny and J. Wang, Computing the basic reproductive numbers for epidemiological models in nonhomogeneous environments, Appl. Math. Comput., 242 (2014), 473–490. |
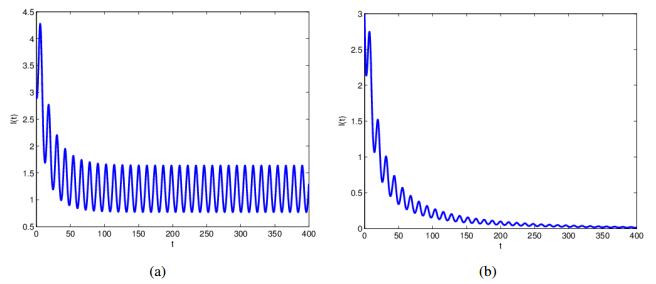
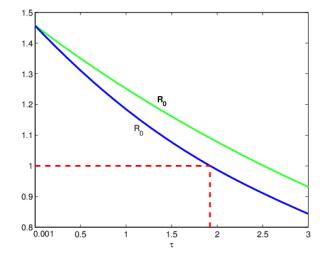
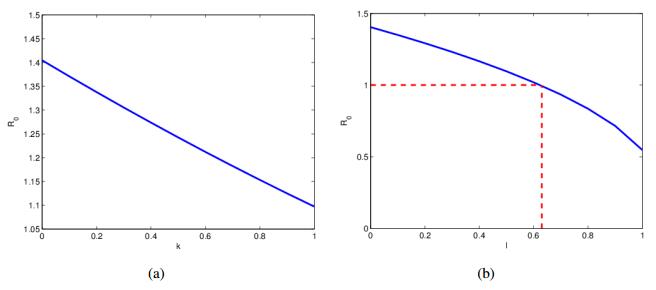
Figures(3)

Junli Liu. Threshold dynamics of a time-delayed hantavirus infection model in periodic environments[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4758-4776. doi: 10.3934/mbe.2019239







 DownLoad:
DownLoad: